-moblog- Having spent this weekend in Heidelberg city at a meeting of the German NGFN project I had the opportunity to listen to an excellent talk of Stephan Beck who works at the Wellcome Trust Sanger Center.
Epigenetics is the connecting link between the rather fixed genome and the variable transcriptome. To start with the end of the talk: Beck predicts for the near future highly parallel SNP, expression and methylation arrays. Although the first methylome has just been published 4 weeks ago by the Arabidopsis community (as with RNAi the plant people again at the forefront) there is still a long way ahead for a first human methylation map.
The latest information may be retrieved from www.epigenome.org, www.epigenome-noe.net, www.epitron.eu,
www.heroic-ip.eu and the German National Methylome Project on chromosome 21 (please google for the link). The methylome is largely an European initiative – the two US epigenome projects do not have any website so far. The network site has some introductory texts; Beck was also refering to a 2006 PLOS paper by Akhtar.
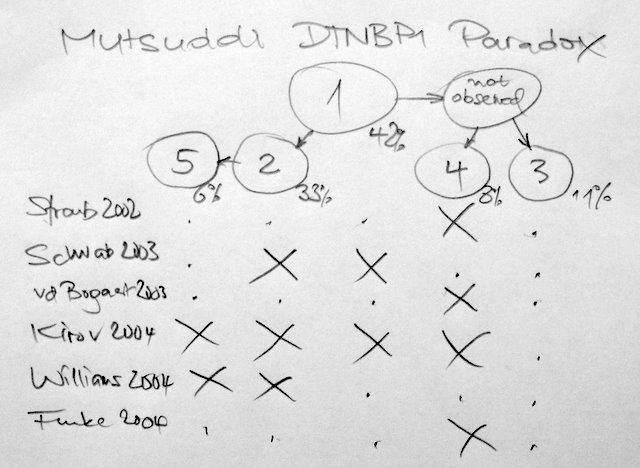
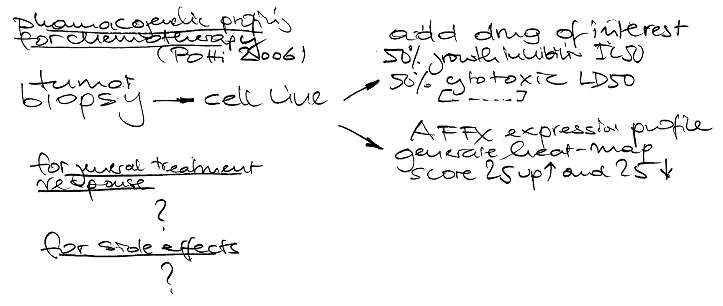
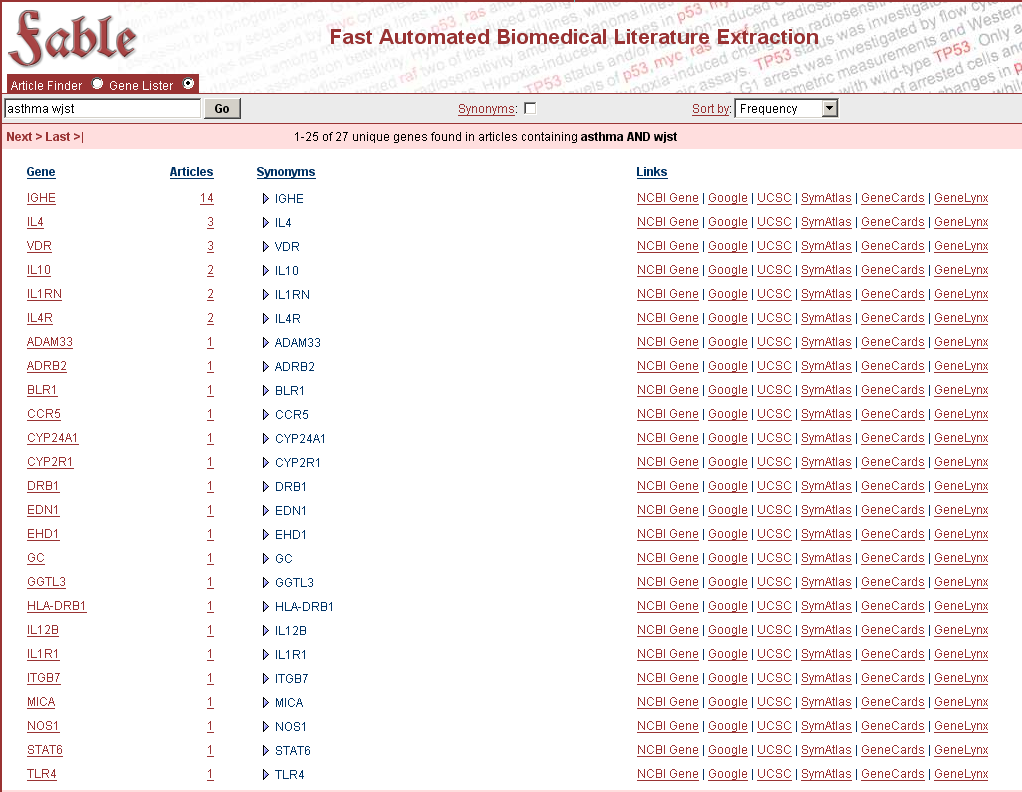
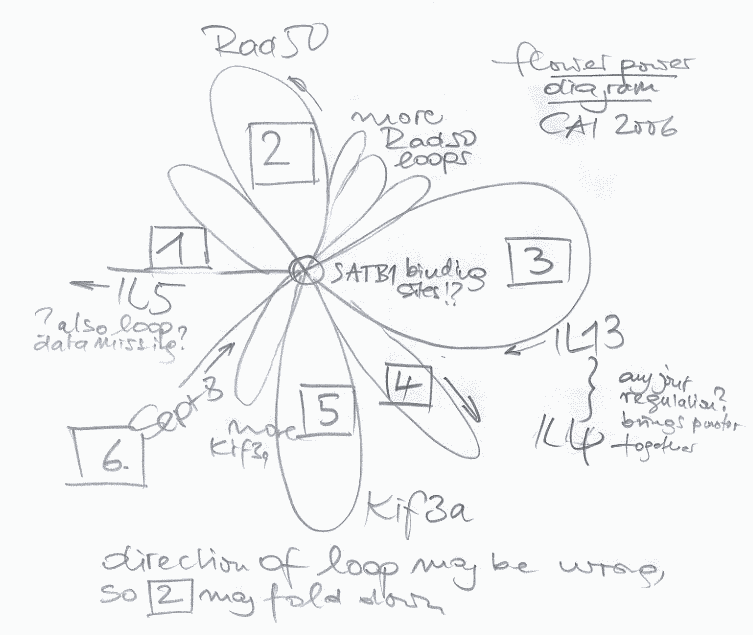
Currently there are 4 human chromosomes under work covering 873 genes (hopefully I captured this correctly as this was a very dense talk). 70% of genes examined so far are either clearly methylated or they are not methylated by testing 12 different tissues. Sperm stands out from all other tissues – which is not unexpected. Tissues originating from the same developmental background have similar methylation patterns – also not unexpected. A preliminary analysis of expression patterns shows that if the 5 prime end is methylated expression is suppressed- also not unexpected.
Fascinating: the colon cells that certainly have a close interaction with the environment do NEITHER show age NOR sex specific differences. Fascinating too: The most frequently methylated regions are ECRs (evolutionary conserved sequences) for whatever reason. Promotor methylation dips around the transcription start sites – from the plots I would say plus and minus 2000bp. Methylation seem to be also conserved between mouse and human tissues while methylation status seems stable over time.
Current bisulfite sequencing is still laborious, expensive and takes quite a long time while immunoprecipitation using MeDIP is getting an alternative. The Sanger people also did a study usinge Nimble(R) gene 50 mers where Ensembl and UCSC will soon have these data for display. Finally, methylation appears in blocks. TagMVPs (your guess is correct, these are tags for methylation variant profiles) construction is straightforward where the estimated 40 million CpG sites will probably be covered by less than 10 percent tagMVP – Haplo epi types are now called hepitypes, yea, yea.

Addendum
Methyl Primer Express® Software â€" is a free software package to simplify and automate the primer design process in methylation experiments. The bisulfite kit is not free ;-)
Addendum
A new textbook and a nice preview
CC-BY-NC Science Surf , accessed 07.08.2026