Notice it never argues with the Society. It can’t – the RCTs are the RCTs. So it just moves the fight somewhere else. The Society shut the door on the low-normal band. This reopens it by calling that same band “deficiency.” The new “deficient” line sits exactly on the floor of the old “insufficiency” band the Society just threw out. Everyone below 50 gets relabelled. In UK Biobank, isn’t that more than half of adults?
The first pages are all rickets, seizures, osteomalacia. Sure, those are real, and I have never said otherwise. But that’s the trick – they open with mis-treated toddlers in Glasgow and close with a cutoff for sixty million people who feel fine.
They half-admit it themselves near the end: efficacy depended on VDR genotype, and “we will need to move away from using serum 25(OH)D thresholds as the only target.” Fine - so why demand a new one?
And then read who’s asking – Frontiers prints it in full:”
SC: Over the last 3 years received consultant fees from the Council for Responsible Nutrition UK CRN UK. ACT NOW Vitamin D's inception meeting was supported by CRN UK, including room hire, subsistence and travel expenses. TH: Over the last 5 years, received consultant fees from Noble Foods and an honorarium from Danone. TH's institution currently receives a sponsored PhD studentship from The Lakes Free Range Egg co. Currently instructed as an expert witness by a professional services firm in a tribunal matter concerning a nutritional supplement company. Trustee of the Nutrition Society charity. CR: Received consultancy fees during 2025 from the European Fruit Juice Association, Yoplait, General Mills, the UK Tea & Infusions Association, Tate & Lyle, Holland & Barrett, the Proprietary Association of GB, BBC Radio Scotland, the British Egg Industry Consortium, INRAE and the University of Reading. She also serves as a board member of Quality Meat Scotland, a public body. MH: Co-holder of two patents on the therapeutic use of vitamin D binding protein; honorarium and royalties as Editor-in-Chief of Feldman and Pike's Vitamin D. The remaining author(s) declared that this work was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The inception meeting paid for by the supplement trade body. Two patents on vitamin D binding protein. Royalties on a textbook.
I’m not saying the rickets isn’t real. But do you understand the urgency? I can only speculate. The ground moved – an earthquake – and an establishment defending its liturgy. Fresh scary NDNS numbers, a government that still hasn’t answered its own 2022 call for evidence, and SACN sitting at 25 as the world’s outlier.
If you are a MD: please treat vitamin D deficiency where you find it, the child kept indoors, the elderly housebound or dark skin in a Northern winter. But don’t turn half the country into patients three weeks after the biggest society on earth said stop that.
Yea, yea. Nay, nay. The rest cometh of the trade associations.
Sowohl das Deutsche Ärzteblatt als auch die Münchner Medizinische Wochenschrift veröffentlichten nun in kurzer Folge methodisch fragwürdige Beiträge zur Vitamin-D-Forschung. Und da sie eine enorme Reichweite in der ärztlichen Fortbildung haben, landet der Vitamin-D-Hype direkt in der Praxis mit den bekannten Folgen: Überdiagnostik, unnötige Supplementierung, wir kennen die Probleme viele Jahre zB hier. Von einer Erwiderung habe ich dennoch abgesehen, da dem Erstautor der Schlusskommentar vorbehalten bleiben sollte.
Fall 1
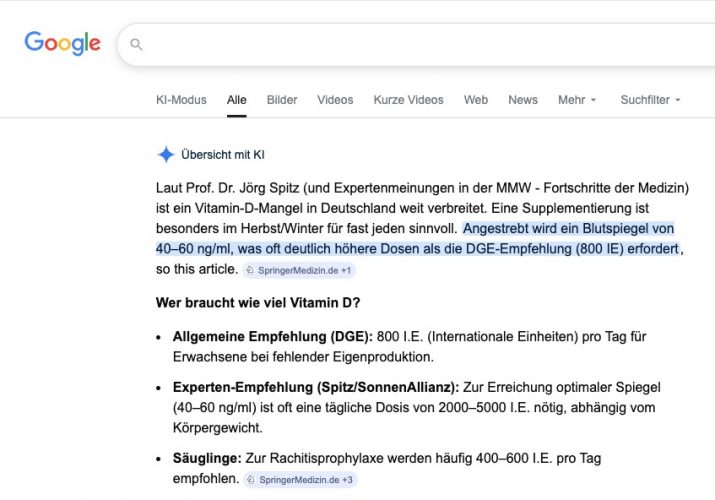
Die MMW druckt eine “Fortbildung”, genauer einen CME-Beitrag "Wer braucht wann wie viel Vitamin D?" eines lange pensionierten fachfremden Arztes (MMW Fortschr Med. 2025; 167 (S3): 76-82). Leider übernimmt die KI den Unsinn nun als Expertenmeinung…
Screenshot Google 26.3.2026
Mein Kommentar zu dem Artikel war
1. Interessenkonflikt
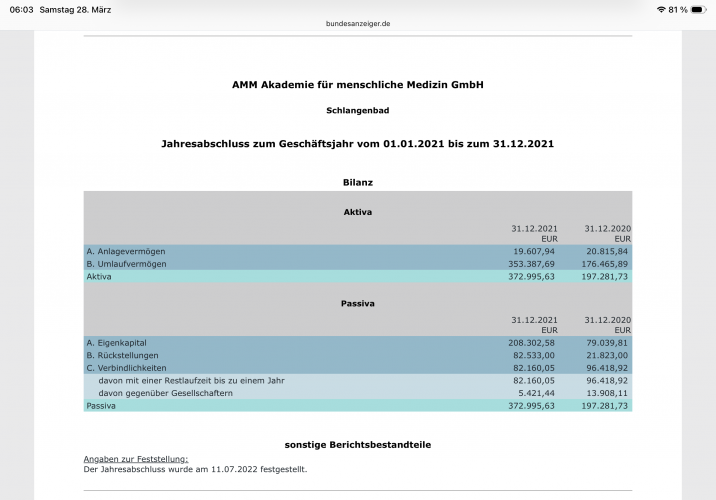
Der Autor gibt "keine Interessenkonflikte" an, betreibt jedoch die Akademie für menschliche Medizin GmbH, die kommerziell Vitamin-D-bezogene Präventionsangebote vertreibt. Diese Verbindung ist gemäß DFG- und ICMJE-Regeln deklarationspflichtig. Medwatch schreibt von 225.000€ Umsatz im Jahr 2019.
2. Epidemiologische Angaben
Die Behauptung, "90 % der Bevölkerung" hätten < 30 ng/ml Vitamin D, stützt sich auf veraltete RKI-Daten (2007-2011). Neuere Erhebungen (DEGS II, 2019) zeigen nur 30-40 % unter 50 nmol/l. → Übertriebene Darstellung eines Mangels.
3. Dosierungsempfehlung
Empfohlene 4 000-6 000 IE/Tag überschreiten den international anerkannten Upper Level von 4 000 IE/Tag (EFSA 2023). Für gesunde Erwachsene fehlt jede Evidenz.
4. Toxizität und "Coimbra-Protokoll"
Die Aussage, Werte bis 150 ng/ml seien unbedenklich, widerspricht Daten zu Hyperkalzämie und Nephrokalzinose. Das "Coimbra-Protokoll" (≥ 100 000 IE/Tag) ist nicht evidenzbasiert und medizinisch riskant.
5. Extra-skelettale Effekte
Im Artikel werden präventive Wirkungen (Demenz, KHK, Krebs, Blutdruck, Diabetes u. a.) behauptet, obwohl große RCTs (VITAL, D2d, ViDA) keinen Nutzen zeigten. Die Darstellung ist selektiv und irreführend.
6. Schwangerschaft
Die genannte 60 %ige Reduktion von Frühgeburten entstammt keiner belastbaren Quelle; die zitierte Pilotstudie (Wagner et al. 2006) weist diese Endpunkte nicht auf.
7. COVID-19-Bezug
Die angeblich 16-fache Mortalitätssteigerung bei Vitamin-D-Mangel beruht auf einem Meinungsartikel, nicht auf einer Originalstudie. Die Darstellung ist faktisch falsch.
8. Fehlende Neutralität im CME-Kontext
Der Beitrag enthält werblich anmutende Aussagen, selektive Quellenwahl (u. a. Masterarbeit Göthel 2020) und unkritische Übernahme von Hypothesen. Damit ist die formale Neutralitätsanforderung der CME-Zertifizierung (§ 4 CME-Richtlinie BÄK) nicht erfüllt.
Der Chefredakteur Cornelius Heyer nimmt auf meine Reklamation zwar die CME Akkreditierung heraus, aber statt einer Korrektur oder Löschung des Beitrages, druckt die MMW einen Leserbrief nach dem Spitz das letzte Wort in der Diskussion hat. Spitz in Bhakdi Manier “Die von mir dargestellten Fakten entsprechen der aktuellen internationalen Forschung – wenn die Leitlinien das nicht tun bitte ich, dies mir nicht anzulasten”.
Screenshot 26.3.2026 Nota bene – Spitz hat nie wissenschaftlich zu dem Thema gearbeitetPassiva der Akademie für menschliche Medizin 2021
Ein beliebiger Leser der Tageszeitung ist damit nun also besser informiert als ein/e Arzt/Ärztin der die MMW abonniert hat.
Fall 2
Das Deutsche Ärzteblatt druckt die Besprechung eines Kongressabstracts “Personalisierte Vitamin D Supplementierung kann das Re-Infarktrisiko halbieren”. Wer nec ist weiss ich nicht, allerdings weiss ich genau, dass es wieder ein Märchen ist das nec hier erzählt. Der Leiter der Medizinisch-Wissenschaftlichen Redaktion des DÄ Christopher Baethge verweist mich an die journalistische Redaktion in Berlin Michael Schmedt, der allerdings auch nach Wochen immer noch nicht geantwortet hat.
New Orleans - Eine Supplementierung mit Vitamin D in individuell titrierter Dosierung kann das Risiko für einen erneuten Herzinfarkt bei vorerkrankten Personen um mehr als die Hälfte reduzieren. Das berichteten Forschende bei den Scientific Sessions 2025 der American Heart Association in New Orleans (Abstract Nr. 4382525).
[...]
Warum ist das so?
Die TARGET-D-Studie wird als Korrektur früherer negativer Vitamin-D-Studien präsentiert, mit der Behauptung, diese seien gescheitert, weil sie Vitamin D nicht "zielgerichtet" titriert hätten. Bei genauer Betrachtung reproduziert TARGET-D jedoch genau jene strukturellen Schwächen, die die Vitamin-D-Outcome-Literatur seit Jahren geplagt haben.
Der entscheidende Befund ist eindeutig: Der vordefinierte primäre Endpunkt, schwere kardiovaskuläre Ereignisse (MACE), wurde in der Intention-to-treat-Analyse nicht signifikant reduziert. Nach elementaren CONSORT-Prinzipien müsste dieses Ergebnis die Interpretation bestimmen. Stattdessen verlagert die Studie den Fokus sofort auf ein einzelnes positives Signal in einer Unterkomponente des kombinierten Endpunkts, den Folge-Myokardinfarkt. Diese Verschiebung rettet die Studie nicht, sondern verschleiert ihr negatives Hauptergebnis. Wenn ein kombinierter Endpunkt scheitert, stellt die selektive Hervorhebung einer einzelnen Komponente keine Evidenz dar, sondern narrative Verzerrung.
Die anschließende Betonung von Per-Protocol-Analysen untergräbt die Aussagekraft weiter. Diese Analysen vergleichen nicht mehr randomisierte Gruppen, sondern Untergruppen, die nachträglich anhand des Erreichens eines willkürlich gewählten Vitamin-D-Schwellenwertes definiert werden. Damit wird nicht mehr die Wirkung einer Intervention geprüft, sondern der Gesundheitszustand einer selektierten, therapietreuen, Responder Population dem einer Restgruppe gegenübergestellt, die überproportional Nicht-Responder, Gebrechliche und Patienten mit höherer Krankheitslast enthält. Der Vitamin-D-Spiegel fungiert hier als Marker guter Gesundheit und Compliance, nicht als kausaler Faktor. Genau dieser Denkfehler hat frühere Beobachtungsstudien diskreditiert, was auch in allen Umbrella Reviews klar herauskam, aber hier innerhalb eines randomisierten Designs erneut eingeführt wird.
Das Ausmaß der postrandomisierenden Selektion macht dieses Problem unübersehbar. Ein erheblicher Teil der dem Vitamin-D-Arm zugewiesenen Patienten wird aus der Per-Protocol-Analyse ausgeschlossen, weil der Zielwert nie erreicht wurde oder nur unvollständige Nachbeobachtung vorlag. Die Konditionierung auf einen postrandomisierten Biomarker zerstört die durch Randomisierung erreichte Vergleichbarkeit der Gruppen. Die daraus resultierenden Effekte sind selektionsgetrieben und nicht kausal interpretierbar.
Auch die biologische Grundannahme der Studie ist schwach fundiert. Der gewählte Zielwert von über 40 ng/ml für 25-Hydroxyvitamin D ist nicht leitlinienbasiert, entspricht nicht der neuesten Literaturund impliziert einen Schwellen- oder Dosis-Wirkungs-Effekt, der in großen randomisierten Studien und genetischen Analysen nicht bestätigt wurde. Indem der Studienerfolg über das Erreichen dieses Zielwerts definiert wird, wird der behauptete Nutzen nicht getestet, sondern implizit vorausgesetzt.
Die Darstellung der Ergebnisse verstärkt diesen Eindruck. Formulierungen wie "klinisch relevante Risikoreduktionen" oder "Reduktion des Myokardinfarktrisikos um mehr als die Hälfte" suggerieren einen kausalen Effekt, den das Studiendesign und die Resultate nicht tragen. Andere Komponenten des kombinierten Endpunkts zeigen keine konsistente Verbesserung, werden jedoch deutlich weniger betont. Der Gesamteindruck ist nicht der einer neutralen Prüfung einer Hypothese, sondern der Versuch, aus einer im Kern negativen Studie ein positives Narrativ zu extrahieren.
Insgesamt liefert TARGET-D keinen belastbaren Beleg dafür, dass eine Vitamin-D-Normalisierung das kardiovaskuläre Risiko nach akutem Koronarsyndrom senkt. Der primäre Endpunkt ist negativ, die sekundären Aussagen beruhen auf selektiver Gewichtung, und die Per-Protocol-Analysen ersetzen Randomisierung durch Adhärenz- und Selektionsmechanismen. Die Studie bestätigt letztlich nur, was die Literatur seit Langem zeigt: Niedrige Vitamin-D-Spiegel korrelieren mit schlechter Gesundheit, ihre Korrektur verändert jedoch harte kardiovaskuläre Endpunkte nicht zuverlässig. Neu an TARGET-D ist nicht die Überwindung dieser Limitationen, sondern ihre methodisch aufwendigere und potenziell irreführende Verpackung.
Die Assistentin der Chefredaktion Monia Jacobs entgegnet sinngemäß dann am 15.4. per Email, die Studie sei auf dem renommierten AHA-Kongress vorgestellt und von der AHA selbst per Pressemitteilung kommuniziert worden – ergo sei das nicht Problem des Ärzteblattes sondern der Kongressveranstalter. Der eigene Beitrag würde die Ergebnisse transparent darstellen, einschließlich des verfehlten primären Endpunkts – klassische narrative Verzerrung. Journalistische Sorgfaltspflicht verlangt, auch die gegenteilige Meinung zu benennen – sofern man denn überhaupt verstanden hat worum es hier geht.
I can’t even remember how many vitamin D studies we did, explaining how this prohormone has been discovered and how stupid guidelines came on to the scene.
And it didn't happen quietly. It wasn't a minor tweak, a footnote, or an incremental update. It was a full reversal of a doctrine that has dominated labs, clinics, public-health brochures, and countless biomarker panels for decades. A classical paper even claimed that 50% of the world population is vitamin D insufficient. For years, we had to live with the tidy triplet:
That middle category "insufficiency" became a diagnosis in itself. It justified mass screening. It justified supplementation campaigns. It justified entire clinical cultures built around chasing numbers. And then 2024 arrived.
Because after reviewing all high-quality randomized trials, the Endocrine Society concluded something truly astonishing:
there is no reliable evidence that people with 25(OH)D levels between 20 and 30 ng/mL derive any clinically meaningful benefit from raising those levels
In fact, the guideline panel found that even below 20-24 ng/mL, evidence for clear benefit is surprisingly weak or uncertain - except perhaps in the very elderly, and even there the benefit didn't map neatly to a threshold. Vitamin D physiology makes the whole "insufficiency" concept biologically dubious, because serum 25(OH)D is only an external storage marker of an intracellular prohormone system - a tank that appears "empty" only in true deficiency like rickets. Let me put that differently: The category of "vitamin D insufficiency," introduced in 2011 and used worldwide, is now considered *scientifically unsupported*. The Society explicitly withdraws it.
That is not merely unusual. In the world of clinical guidelines, this is as close as you get to a scientific earthquake. Why did they withdraw it? Because the evidence never really showed what everyone assumed.
The new communication explains the problem with striking clarity:
1. Observational associations misled us.
Many early threshold claims came from correlations - low vitamin D and higher PTH, low vitamin D and lower bone density, etc. But none of this proved causality, and much of it turned out to be non-informative once RCTs were performed.
2. Surrogate markers were overinterpreted.
Calcium absorption, PTH suppression, even bone mineral density - these are *indirect* signals. They don't automatically translate into fewer fractures, fewer falls, fewer infections, or longer life. And when RCTs finally tested real outcomes, the expected clinical benefits simply weren't there.
3. Large RCTs showed no special benefit in "low-normal" ranges.
VITAL - one of the biggest vitamin D trials ever - found no difference in fractures even in participants below 24 ng/mL, and even those below 12 ng/mL did not exhibit the dramatic benefit everyone predicted (though the subgroup was very small).
4. Across thousands of participants aged 50-74, supplementation beyond the RDA made essentially no difference - including in those below the supposed thresholds.
The forest plots in the guideline communication make this visually obvious: the <20-24 ng/mL subgroups almost never differ from the overall population in any meaningful direction. (See page 5 of the document: identical risk-change estimates for falls, fractures, cancer, CVD, etc.)
We rarely see a major medical society openly dismantle one of its own most influential guidelines - not because of scandal, not because of politics, but because the evidence finally matured and said: we were wrong. And they didn't hedge. They didn't massage the language. They called the new stance what it is: epistemic humility.
Jörg Spitz ist wohl nicht der geeignete Experte für dieses Thema – es sei denn, als würde man auch Wolfgang Wodarg, Michael Meyen, Sucharit Bhakdi oder Stefan Homburg zu COVID befragen wollen.
Der Autor erklärt, dass er sich bei der Erstellung des Beitrags von keinen wirtschaftlichen Interessen leiten ließ. Er legt folgende potenzielle Interessenkonflikte offen: keine.
Der Verlag erklärt, dass die inhaltliche Qualität des Beitrags durch zwei unabhängige Gutachten bestätigt wurde. Werbung in dieser Zeitschriftenausgabe hat keinen Bezug zur CME-Fortbildung. Der Verlag garantiert, dass die CME-Fortbildung sowie die CME-Fragen frei sind von werblichen Aussagen und keinerlei Produktempfehlungen enthalten.
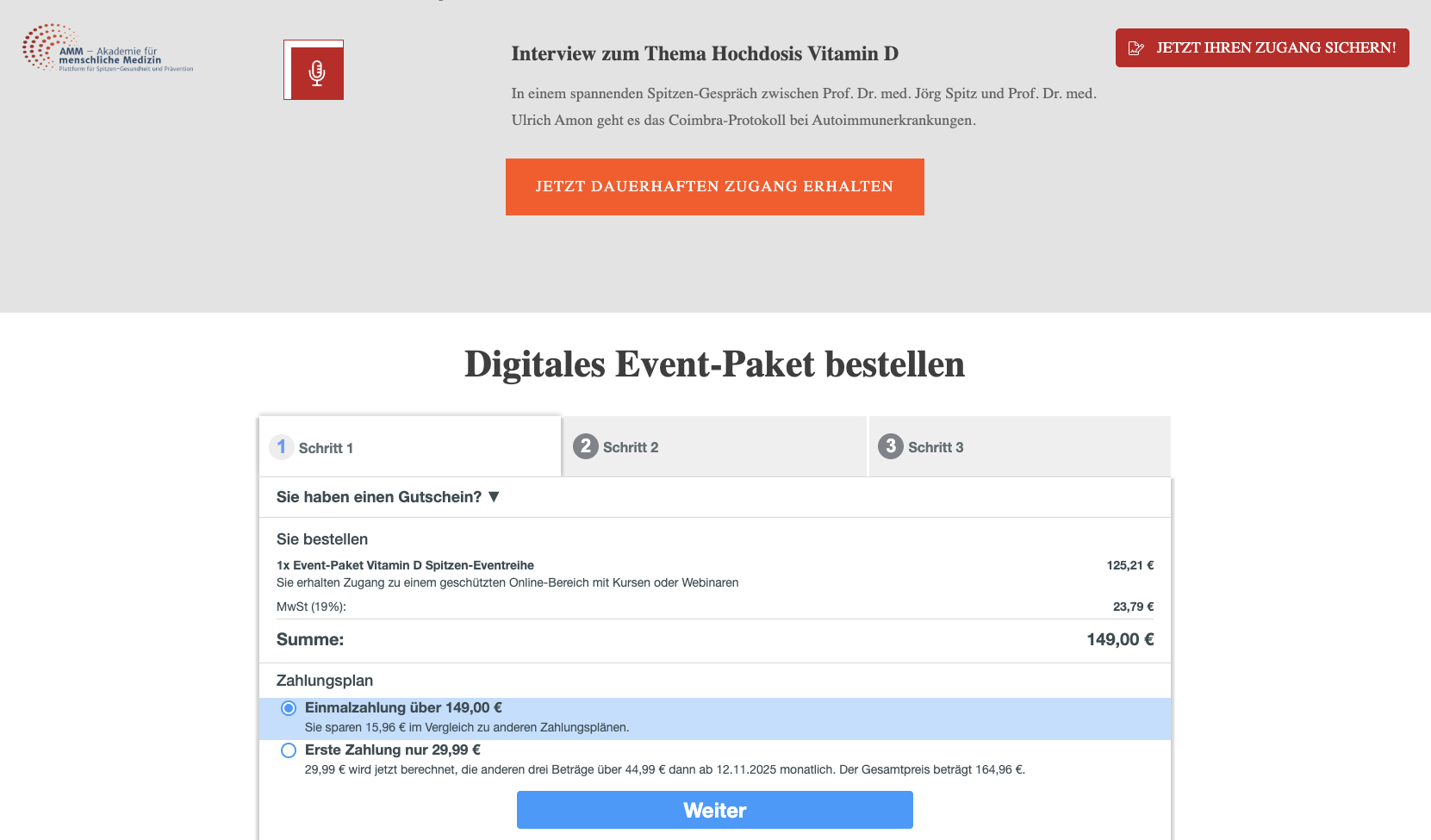
Gleichzeitig vertreibt Spitz aber sein “digitales Event Paket” online zur umstrittenen Hochdosis Vitamin D Therapie für 149€.
Ich hätte das nicht vermutet, aber Vitamin scheint doch ein lukratives Geschäftsmodell zu sein auch wenn man mit der Substanz selbst nichts verdienen kann. So hat medwatch schon vor längerer Zeit festgestellt
Hinter der Seite steckt der Nuklearmediziner, Buchautor und selbsternannte Präventionsexperte Jörg Spitz. Und: Vitamin D scheint eines seiner Lieblingsthemen zu sein, so hat er bereits fünf Ratgeber zu dem Vitamin veröffentlicht. Zudem betreibt er ein ganzes Netzwerk aus verschiedenen Internetseiten, die sich allesamt um Vitamin D und weitere Nahrungsergänzungsmittel drehen, etwa die Akademie für menschlichen Medizin (AMM). Die GmbH machte in 2019 einen Umsatz von 225.000 Euro - Spitz ist ihr alleiniger Mitarbeiter.
Und die Referenten der spitzen-praevention.com ? Ein echtes Panoptikum , die meisten ohne wissenschaftliche Qualifikation, dafür aber mit Missionseifer.
Yep there was some hype about 100 years ago, soon after the discovery of vitamin D by Windaus – where vitamin D / cod liver oil was used to treat tuberculosis.
Also vitamin A (also contained in high doses in cod liver oil) was used in the pre-antibiotic era as anti-infective therapy while it has lost its role being used only as some supportive add-on after during measles infection. Reliable prevention of measles by vaccination should not be confused with uncertain supportive therapy.
But what about vitamin D supplementation and cancer? A new study in the Annals last week show
7% reduction in cancer mortality was observed after a median cumulative follow-up of 22.3 years (1817 vs. 1943 deaths; hazard ratio [HR], 0.93 [95% CI, 0.87 to 0.99]), along with a 6% increase in CVD mortality (2621 vs. 2420 deaths; HR, 1.06 [CI, 1.01 to 1.12]).
As both risks are of borderline significance and we don’t trust P values alone, my conclusion is: vitamin D supplementation doesn’t prevent cancer.
In addition to some programming questions, I have prompted several LLMs for a summary of my longstanding research question “vitamin D supplementation in the newborn period and later allergy”. Here is the elicit response:
Research on vitamin D supplementation in infancy and its relationship to later allergy development has yielded mixed results. Some studies suggest that higher vitamin D intake during the first year of life may increase the risk of atopic conditions later in childhood (Bäck et al., 2009; Hyppönen et al., 2004). These findings are supported by animal studies and epidemiological surveys linking early vitamin D supplementation to allergy development (Wjst, 2006). However, a randomized controlled trial found that high-dose vitamin D supplementation did not prevent allergic sensitization or diseases in infants compared to standard dosage, except for an increased risk of milk allergy in the high-dose group (Rosendahl et al., 2019). Interestingly, this study also observed that infants with high cord blood vitamin D levels had a higher risk of food allergen sensitization. These conflicting results highlight the complex relationship between vitamin D and allergy development, suggesting that both insufficient and excessive vitamin D levels may potentially influence allergic outcomes.
The limitation introduced by “however” is wrong – the whole story is not really conflicting if you read my earlier paper on a paradoxical reaction. I have been a reviewer of the Rosendahl paper and asked them to change their misleading spin. It is basically a triple blind study (the authors believed the opposite fact eg that vitamin D supplementation prevents while it induces allergy). Unfortunately, as Rosendahl et al. did not respond to my complaint, we can now clearly see how the bias of the authors is being injected into the LLM.
I dont know why there is still the discussion of any genuine understanding of LLMs as there is none, see Binz et al.
even though LLMs often achieve state-of-the-art performance on existing benchmarks, it remains debated whether this reflects genuine understanding, or whether they are merely acting like stochastic parrots […] When deciding whether to use an LLM, it is important to recognize that LLMs are simply models of word form distributions extracted from text-not models of the information that people might get from reading that text
Anyway, the workflow suggested by inkenai – putting these PDFs now into NotebookML for further analysis – is excellent.
A new paper from Bristol discusses the recent explosion of low-quality two-sample Mendelian randomization studies and offers a cure.
We advise editors to simply reject papers that only report 2SMR findings, with no additional supporting evidence. For reviewers receiving such papers, we provide a template for rejection.
We know it for ages (1985) that 1,25-Dihydroxyvitamin D3 suppresses human T helper cells by blocking interleukin 2 (IL 2) production. The immune effects were our main argument 2001 against oral vitamin D supplementation of newborns as it may induce later allergy . It is now funny to find now a clinical trial that tests low-dose interleukin-2 in birch pollen allergy.
to ask also chatGPT consensus some of my lifelong research questions
nothing wrong here but clearly not weighted at all. Maybe the consensus app need to learn also rules of meta-analysis and evidence based research in particular when attention to the details is decisive for epidemiology, immunology and clinical studies.
Ich hatte direkt nach der Wende in Halle und Leipzig die erste Ost-West Allergiestudie* durchgeführt und dabei die verblüffende Entdeckung gemacht, dass es im Osten nicht mal halb so häufig Allergien gab.
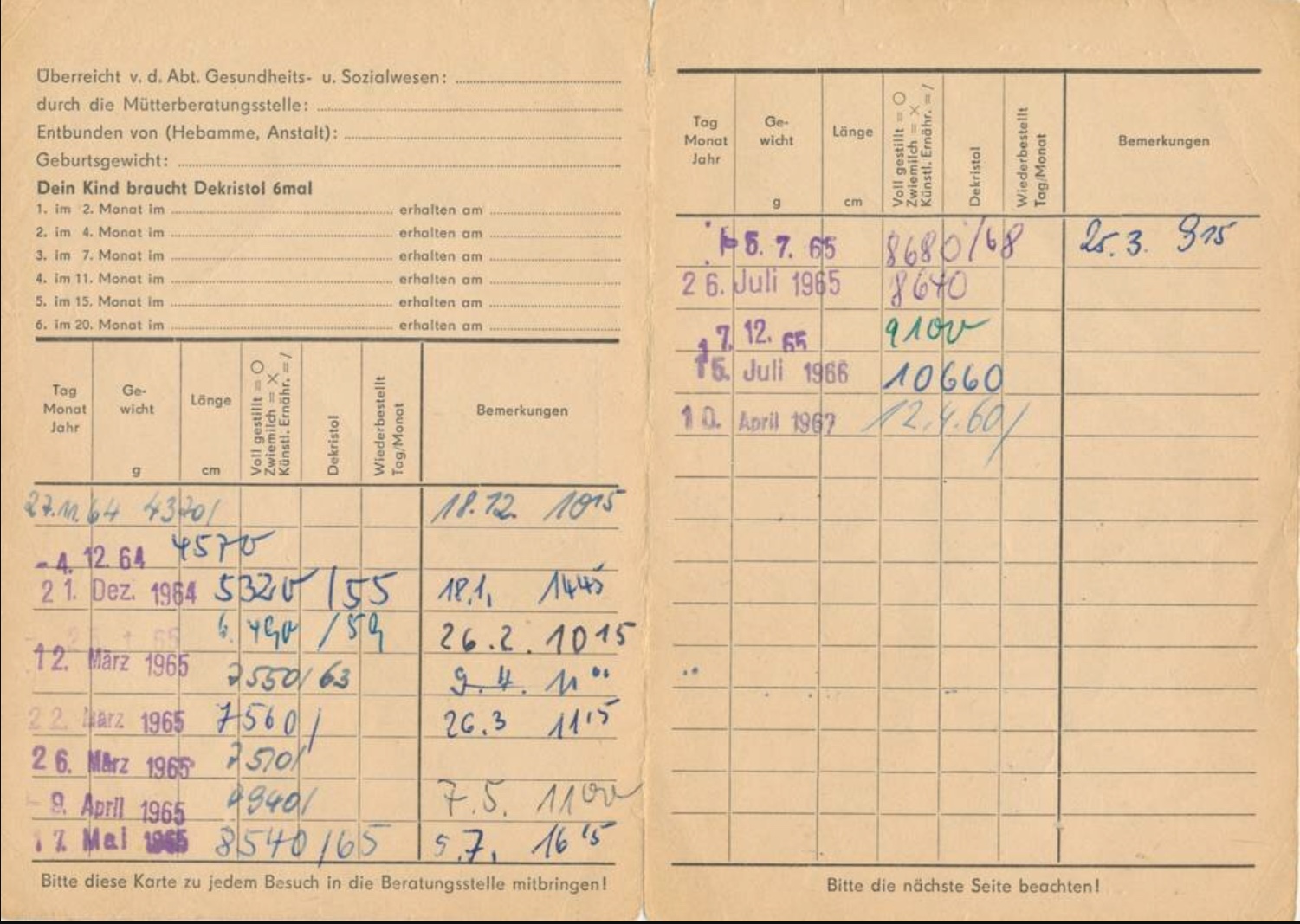
Der Grund dafür war uns zunächst nicht klar, aber dann stelle sich doch bald heraus, daß es wohl die unterschiedliche Vitamin D Prophylaxe war – im Westen gab es tägliche niedrige Dosen ab der ersten Lebenswoche, im Osten wurde ab dem 2. Monat wenig hohe hohe Einzeldosen verabreicht. Wenn Kinder im Osten krank waren, dann fiel auch immer wieder die eine oder andere Dosis aus, auch wurde das Schema nach meiner Recherche nicht immer komplett durchgezogen. Leider haben wir uns damals aber nur die Impfpässe angesehen, nicht aber die im Nachinein wichtigeren Wiegekarten.
DDR Wiegekarte. Quelle https://www.ddr-museum.de/de/objects/1011262Westdeutsches Untersuchungsheft U2. Quelle https://www.g-ba.de/downloads/83-691-452/2017-05-11_GBA_Kinderuntersuchungsheft_Web-WZ.pdf
Die Befürchtung der Professoren Mai und Beuren in dem alten SPIEGEL Artikel über Vitamin D Nebenwirkungen haben sich zum Glück nicht bewahrheitet. Dafür aber stellte sich dann aber eine unerwünschte immunologische Wirkung heraus die damals noch nicht bekannt war.
Mit dem aus England bzw Amerika importierten Schema zur Supplementierung stiegen die Allergien an. In der BRD war das ab den frühen 60er Jahren , wie überhaupt die englischsprachigen Länder immer die höchsten Allergiehäufigkeiten hatten, da sie wegen der häufigen Rachitis auch viel konsequenter supplementierten (die Rachitis hiess früher einmal auch “englische Krankheit”).
Der Mechanismus der Allergieentstehung ist dabei nur teilweise aufgeklärt: Vitamin D ist jedenfalls immunsuppressiv mit vielfacher Auswirkung auf B und T Zellen was seit dem Nachweis des Vitamin D Rezeptors auf diesen Immunzellen wissen. Die Supplementierung stört offensichtlich die initiale Klassifikation ob ein Protein harmlos oder allergen ist.
Warum es aber auch schon 1989 Allergien im Osten gab? Nun, es war ja keine Vitamin D freie Zone, offensichtlich sensibilisiert man sich auch noch ausserhalb des überkritischen Intervals in den ersten Lebenswochen.
It seems that I missed an interesting 2017 paper that looked for disease-associated SNPs in canonical DR3 motifs. Only 7 out of 211 traits showed significant hits, one of these was self-reported allergy. When annotating these SNPs, there are only two genes: LINC00299 and TLR1
So are TLR1 & LINC00299 variant carriers more susceptible to vitamin D induced allergy?
LINC00299 (Long Intergenic Non-Protein Coding RNA 299) is a RNA Gene of largely unknown function, associated so far with allergy only on a genetic level in Framingham, href=”https://pubmed.ncbi.nlm.nih.gov/23817569/”>23andme and other studies. We don’t know so much here, the function of the long non coding RNAs
depends on subcellular localization. Depending on their niche, they specifically interact with DNA, RNA, and proteins and modify chromatin function, regulate transcription at various stages, forms nuclear condensation bodies and nucleolar organization. lncRNAs may also change the stability and translation of cytoplasmic mRNAs and hamper signaling pathways. Thus, lncRNAs affect the physio-pathological states and lead to the development of various disorders, immune responses, and cancer.
The TLR1 genetic association is found by many genetic studies, while the clinical association is probably more by an infectious origin. TLR1 is a pattern recognition receptor with a specificity for gram-positive bacteria and also included in my forthcoming exome paper as a protective factor for asthma/allergy. And we are also close to my earlier review of vitamin D, the microbiome and allergy…
Does any co-infection response during first vitamin D exposure influence allergic sensitisation? There are indeed some hints of an short-lived effect of lung group 2 innate lymphoid cells (ILC2s)
Laboratory mice cohoused for 2 weeks had impaired ILC2 responses and reduced lung eosinophilia to intranasal allergens, whereas these responses were restored in mice cohoused for ≥2 months. … These findings suggest that ILC2s respond dynamically to environmental cues and that microbial exposures do not control long-term desensitization of innate type 2 responses to allergens.
I have criticized in the past the milk industry sponsored research that is now operating at a similar scale like research funded by cigarette industry [1]. And well, it seems that I am not alone here when reading a new CEA paper “Are paediatric allergy services promoting or harming public health?”
There were increasing and excessive sales of specialized formula designed for infants with milk allergy-either extensively hydrolysed or amino acid formula. These represented 7.6% of formula sold in 2019 … Although pediatric allergy services cannot be held solely to blame for whole population trends, there is evidence that the clinical guidance used in pediatric allergy clinics might promote or exacerbate these trends. Moreover, many allergy, gastroenterology and pediatric societies and professionals routinely flaunt public health guidance from the World Health Organization by continuing to accept funding from formula companies, despite World Health Assembly resolutions that such conflictive relationships should be avoided.
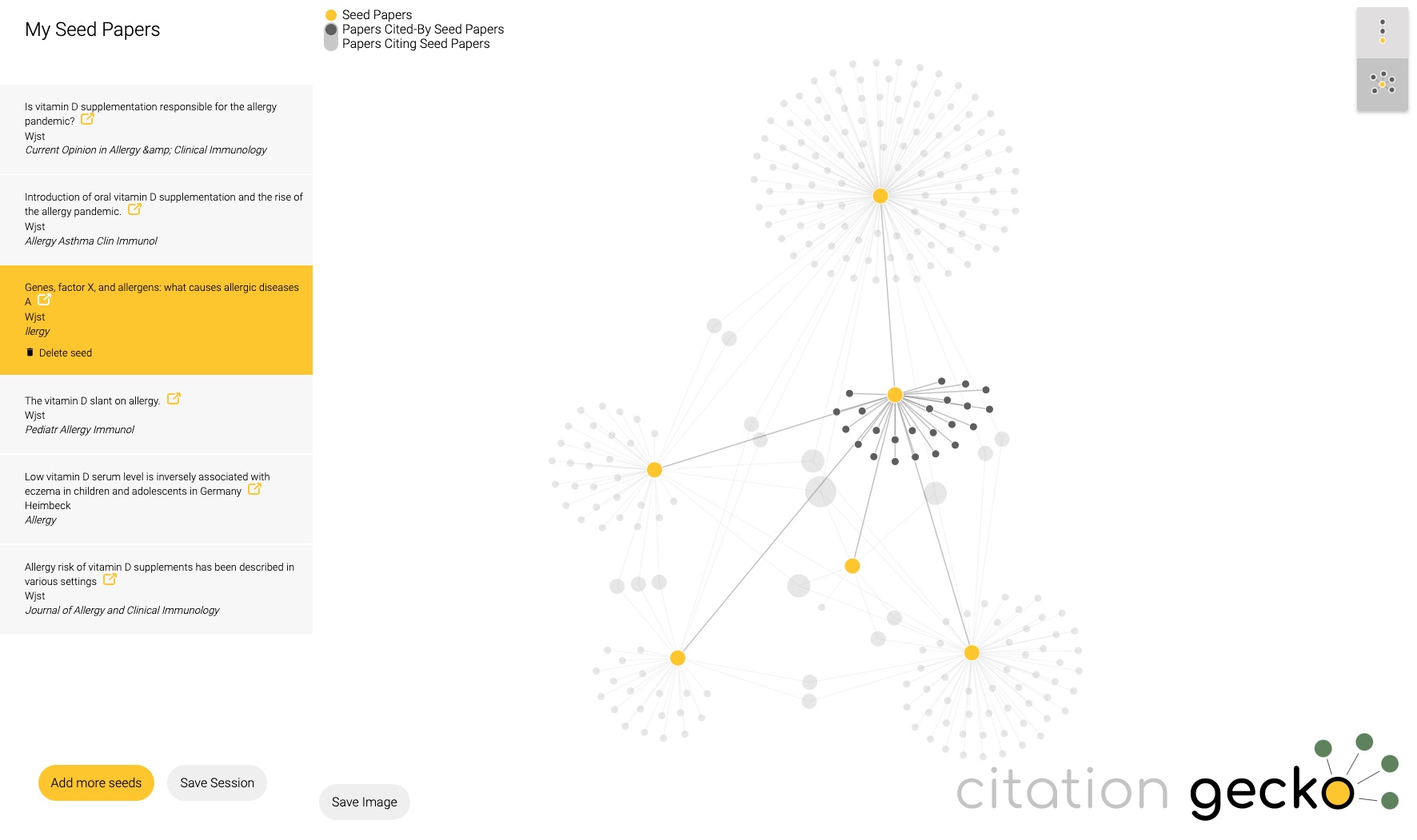
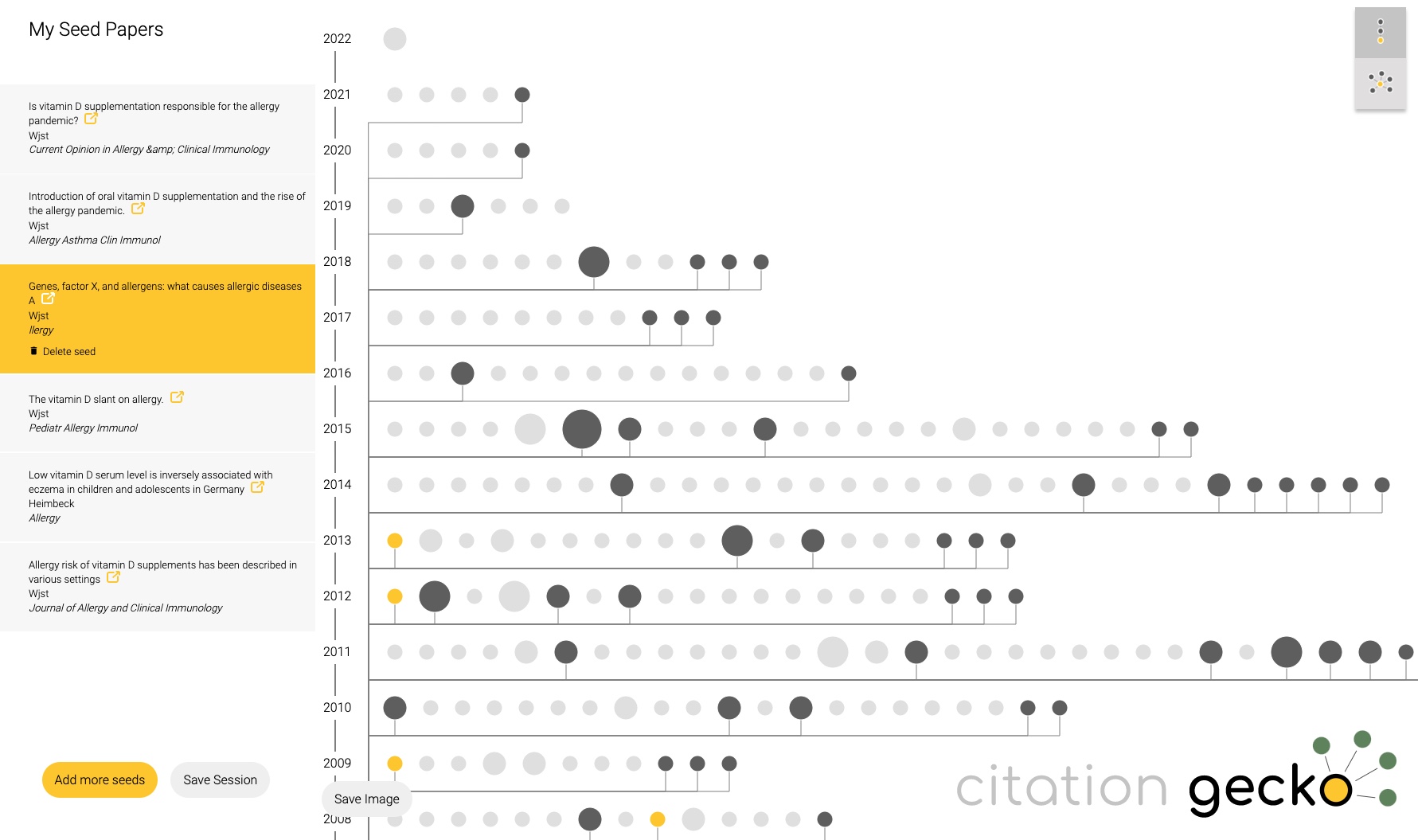
Just tried citationgecko.com on a topic that I have been working on for 2 decades. It will find rather quickly the source paper, much faster than reading through all of it. Unfortunately reviews are rated to be more influential than original data as Citation Gecko picks articles with many references.
Here is the ultimate and comprehensive summary written by two of the best vitamin D experts – Martineau and Cantorna
… lack of a consistent protective signal from RCTs reporting so far is reflected by the absence of any recommendation relating to prophylactic or therapeutic use of vitamin D for COVID-19 in guidelines from national or international bodies … If vitamin D does have favourable immunomodulatory effects in COVID-19, then demonstrating a meaningful clinical benefit of supplementation over existing standards of care is likely to become increasingly challenging, as ever more effective pharmacological therapies and vaccines emerge.
This year vitamin D nonsense information of the year comes by a plant biologist at Rothamsted Research in Harpenden, UK who wants to block a tomatoe conversion pathway leading to an “accumulation of the vitamin D precursor” (which reminds me of the 1930 attempts of mushroom irradiation to generate high amounts of vitamin D2).
Martin and her colleagues plan to study this, and have been given permission to grow their gene-edited tomatoes in fields. The team also hopes to measure the impact of outdoor UV light exposure on conversion of provitamin D3 to vitamin D3 in the plants' leaves and fruit …You can only edit what you understand," he says. "And it's only because we understand the biochemistry that we're able to make those sorts of interventions."
Vitamin D supplements are clearly over rated according to a recent BMJ paper. No doubts that it is a bad idea to supplement an uncontrolled amount of an pro-hormone by eating tomatoes.
Fortunately with 0.2 μg/g or 8 IU D3/g dried tomatoe we are below toxicity levels.